Personality, motivation, and performance
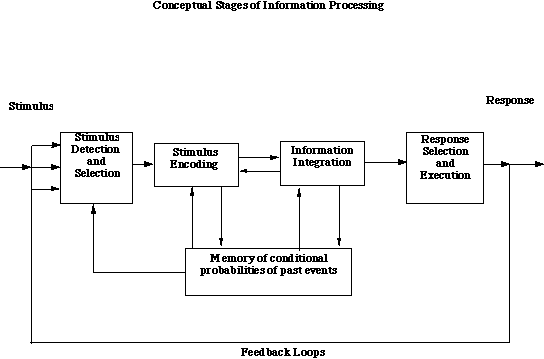
3) A conceptual organization of the stages of processing that are affected by individual differences in motivation. Environmental inputs are first detected, then encoded, stored, and integrated with prior expectancies before responses are selected and executed. Behavioral acts, through feedback, lead to new environmental input. Storage and retrieval processes are represented as arrows to and from the memory system.
Motivation affects each of these stages. In terms of tasks we have examined, we believe that vigilance-like tasks relate to the detection and response stages and are affected by variations in arousal; individual differences in the learning of affectively valenced material occur at the encoding stage and are related to differential sensitivities to rewards and punishments; memory storage and retrieval and the effect of retention interval are affected by variations in arousal: arousal facilitates storage but hinders retrieval; and the information integration stage is curvilinearly related to arousal because it reflects two components--a beneficial effect due to the speed of input and a detrimental effect due to unavailability of recent events.
On a larger time scale, as the information processing loop continues to be executed, resources vary in their availability and in their allocation. Knowledge structures in memory change, affective reactions to the outcomes bias expectancies of future reinforcement and strategic decision processes are used. The encoding of environmental demands reflect differences in biological sensitivities to cues for rewards and punishment (Gray, 1981) as well as the prior contents of memory. Emotional reactions to feedback reflect the interaction of expectancies and outcomes. Positive affective states result from reward following expectancies of reward or non-punishment following expectancies of punishment. Negative affective states result from punishment following expectancies of reward and from punishment following expectancies of punishment (Rolls,1990). Positive affect facilitates approach behavior, negative affect facilitates avoidance behavior. Approach and avoidance tendencies are mutually inhibitory. Increased arousal facilitates the detection and storage of information as well as the execution of the dominant response tendency. This leads to a much more complex model (Figure 4), but one that is probably necessary if the interdependent effects of cognitive and affective processes are to be understood. This model is an attempt to sketch out the systems that are involved in actively processing valenced information in an ongoing system responding to environmental demands and environmental reinforcements[9].
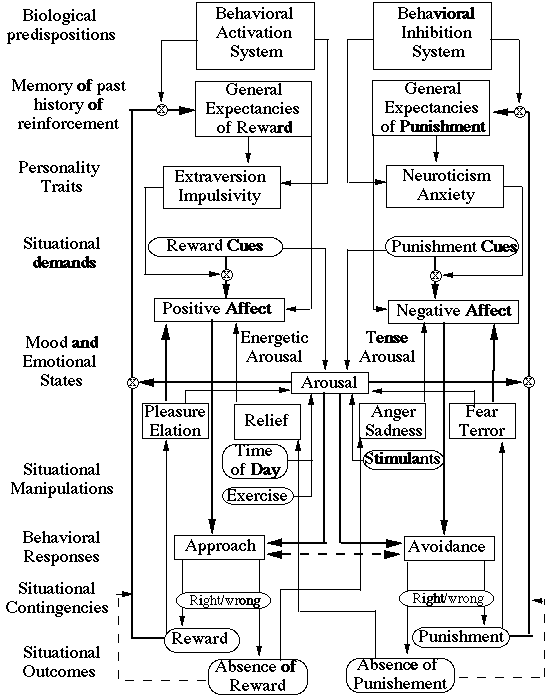
4) Affective and cognitive reactions as part of an ongoing behavioral system. The encoding of environmental demands reflect differences in biological sensitivities to cues for rewards and punishment (Gray, 1981) as well as the prior contents of memory. Reactions to feedback reflect the interaction of expectancies and outcomes. Positive affective states result from reward following expectancies of reward or non-punishment following expectancies of punishment. Negative affective states result from punishment following expectancies of reward and from punishment following expectancies of punishment (Rolls,1990). Positive affect facilitates approach behavior, negative affect facilitates avoidance behavior. Approach and avoidance tendencies are mutually inhibitory. Increased arousal facilitates the detection and storage of information as well as the execution of the dominant response. Adapted from Clark and Watson (1991), Gray (1981), Larsen (1991), Rolls (1990), and Thayer (1989).
In the following section I discuss the immediate motivational effects upon performance of various combinations of individual differences in personality and situational stressors. In the final section I suggest how an adequate theory of individual differences and cognitive performance needs to examine motivational effects on the stages of processing as well as consider the larger temporal variations in affect, cognition and behavior that occur as the information processing loop continues over time.
Personality, vigilance and continuous performance
Differences in the ability to sustain performance across time have been noticed in dogs, sonar operators, train engineers, and faculty listening to colloquia. What is particularly interesting to those interested in coherent descriptions of personality is that several of the basic dimensions of personality are related to performance decrements across time.After an extensive discussion demonstrating that performance decrements generalize across several types of continuous performance tasks, Broadbent (1958) presented evidence suggesting that extraverts were more likely to show such decrements than were introverts. By 1971, the evidence supporting this position was much stronger. Extravert performance deteriorates more rapidly in terms of detecting infrequent signals (Bakan, Belton & Toth, 1963; Keister and McLaughlin, 1972), in terms of variability and speed of continuous reaction time (Thackray, Jones & Touchstone, 1974), and in the ability to stay awake on long distance drives (Fagerström & Lisper, 1977).
Matthews (1989) and Matthews, Davies and Lees (1990) have shown that this decrement in performance can occur very rapidly and that self reported high arousal is associated with the ability to maintain performance. They used a rapidly paced discrimination task introduced by Neuchterlein, Parasuraman and Jiang (1983) with an inter stimulus interval (ISI) of 1 second and a priority stimulus frequency of 25% (i.e. a response was required on the average every 4 seconds) with two levels of stimulus degradation. For the degraded stimuli, performance of low aroused subjects deteriorated within 12 minutes, but did not for high aroused subjects. Neither Neuchterlein et al. nor Matthews et al. found a decrement on this task with non-degraded stimuli. Neuchterlein et al. interpret the detection of the degraded stimuli as requiring substantial "effortful" processing as compared to the non-degraded stimuli which may detected in an "automatic fashion". Matthews et al. argue that the degraded stimuli lead to the kind of resource limited attention task that Humphreys and Revelle (1984) suggest should benefit from high arousal.
Impulsivity at the adult level has frequently been claimed to be related to the impulsivity associated with hyperactivity or what has come to be called Attention Deficit Disorder (ADD) with (or without) Hyperactivity. ADD children are particularly susceptible to decrements on continuous performance tasks. Sergeant and van der Meere (1990) have reviewed the application of Sanders' model of energetic effects on reaction time to the case of individual differences associated with attention deficit disorders. Their review is an excellent example of the wealth of information that comes from combining sophisticated experimental procedures with the study of important individual differences.
Revelle, Rosenberg & Anderson (in preparation) have recently completed three studies with an even simpler task than the Neuchterlein et al. task, but one that still shows pronounced decrements within a few minutes. Because of our interest in the dynamics of behavior, we examined performance as a function of time on task. The task we used (variable fore-period reaction time with an inter stimulus interval of 1-11 seconds) lasts for just a few minutes (12-15) and is typical of the demands placed upon subjects doing many monotonous real world (or experimental) tasks. The subjects task is to respond as rapidly as possible whenever a series of X's appears on the monitor of a computer. The targets remain until the subject responds. The fastest reaction times of our subjects tend to be of the order of 220-250 msec, with most responses being less than 400 msec. We discard all trials in which the subject took more than 1000 msecs to respond, although we have observed at least one subject who was taking 7-8 seconds on some trials. That is, our task succeeds in putting some subjects to sleep. More objectively, self reports of energetic arousal decay reliably across the 12 minutes of the task.
We have done three studies with this task. The first examined the effects of impulsivity, anxiety (neuroticism) and time of day (0900 versus 1930 hours) the second added caffeine as a factor; the third was run just in the morning and examined the effects of an incentive (half of the subjects were offered $10 if they could score in the top 33% of the subjects, the other half were not told about the incentive). Dependent measures were simple reaction time, as well as the change in reaction time as a function of trials.
When the results from all three studies are compared they clearly show a difference between the effects of (caffeine induced or diurnally varying) arousal versus (monetary incentive induced) effort. Although both arousal and effort manipulations improve performance, only the arousal manipulation was able to sustain performance. The change across time clearly demonstrated the effects of arousal as well as impulsivity and neuroticism. Impulsivity was positively correlated with decay of RT in the morning but negatively in the evening, and high neurotics were unable to maintain their performance from the first to the last part of the experiment. These results bring to mind Broadbent's (1971) two levels of control. For although effort facilitated reaction time (Broadbent's lower level) arousal facilitated the long term maintenance of reaction time (Broadbent's higher level).
Personality and non vigilance increments and decrements
Learning valenced material. Humans as well as other animate organisms need to learn sources of reward and punishment within their environment to survive. This fundamental observation has long been ignored by many cognitive theorists concerned with memory. Although a great deal of research on human learning has been done on affectively neutral material (e.g., nonsense syllables), much of the animal learning literature has examined the effects of rewards and punishments upon learning. Jeffrey Gray (1972, 1982) has generalized from an animal model of rat learning to propose a neuropsychological basis of anxiety and to propose a revision of Hans Eysenck's theory of introversion-extraversion and neuroticism. In brief, Gray has proposed that individuals differ in their sensitivities to cues for reward and for cues for punishment. Furthermore, Gray associates the sensitivity to cues for reward with a behavioral activation system (BAS) and the sensitivity to cues for punishment with a behavioral inhibition system (BIS). He associates impulsivity with the BAS, anxiety with the BIS.The evidence for this hypothesis is mixed. Richard Zinbarg and I have shown that when subjects learn a go-no go discrimination task to achieve rewards or to avoid punishments, impulsivity interacts with anxiety to affect rates of learning (Zinbarg and Revelle, 1989). High impulsives who are low on anxiety rapidly learn to make responses to achieve rewards but have difficulty learning to inhibit responses in order to avoid punishment. Highly anxious subjects who are also less impulsive rapidly learn to inhibit their responses in order to avoid punishment. High anxiety when combined with high impulsivity leads to poorer learning, as does low anxiety and low impulsivity. Further support for Gray's model comes from work of Fowles (1980, 1987) and Newman and his associates (Nichols and Newman, 1986; Newman, 1987). Failures to support Gray's hypothesis have been reported by Diaz, Gray & Pickering, (1991) and Pickering (1991).
Kathy Nugent and I extended Gray's model and examined the effect of affective manipulations on the interpretation of stimuli and resulting effects upon memory (Nugent and Revelle, 1991). We examined whether variations in affect (situationally induced by positive and negative feedback) or stable personality traits (impulsivity and neuroticism) affect the memory for neutral stimuli. The results are partly consistent with Gray's model, in that the high impulsives were more likely to remember words following reward rather than punishment, but were inconsistent in that the low impulsives remembered words better following punishment rather than reward (rather than the predicted no effect) and there was no effect of anxiety (we had predicted that more anxious subjects would have better memory for stimuli followed by punishment).
Immediate and delayed retrieval. Honey bees as well as humans need to learn affectively important information and can not afford to waste cognitive resources on trivia. James McGaugh (1990) has reviewed evidence that stimulation following a particular cue enhances the long term memory for that cue. Debra Loftus and I have reviewed 25 years of findings showing that a variety of arousal inductions and measures interact with retention interval to affect memory (Revelle and Loftus, 1990, in press). Experiments using a surprising number of manipulations and measures of arousal have shown similar results: high arousal at learning inhibits immediate retrieval of the information presented but facilitates later recall of that information. Whether this is due to different effects on different stores, or to an arousal induced decrement at retrieval, or to some other explanation remains uncertain. What is certain, however, is that a consideration of individual differences is important. Puchalski (1988) replicated earlier work by Folkard, Monk, Bradbury & Rosenthal (1977) on the effects of time of day on immediate versus delayed retention and found that the pattern reverses for high and low impulsives. Immediate memory of high impulsives was superior in the morning to the afternoon, although recall after one week was superior for information learned in the afternoon rather than the morning. This was essentially Folkard's finding. However, for low impulsives, immediate memory was better in the afternoon than in the morning and delayed recall was equal for information learned at both times of day. Loftus (1990) found that impulsivity and self reported arousal interacted with retention interval to affect the probability of recall. Some of the confusion relating the effects of mood to memory is likely due to ignoring these relationships between individual differences in arousal and the effect of retention interval.
Complex tasks When information needs to be integrated and complex decisions need to be made, there seems to be an optimal level of arousal. Performance on complex reasoning tasks similar to the Graduate Record Exam is an interactive effect of impulsivity, caffeine, and time of day. Specifically, the performance of individuals thought to be less aroused (e.g. high impulsives in the morning, low impulsives in the evening) is facilitated by increases in arousal (e.g. caffeine) while that of individuals thought to be more aroused (e.g. low impulsives in the morning, high impulsives in the evening) is actually hindered (Revelle, Humphreys, Simon, and Gilliland, 1980). This result is large and is replicable. Matthews (1985) has found a similar pattern of results for extraversion and self reported arousal. Although Revelle et al., (1980) suggested that their pattern of results was consistent with an inverted U relationship between arousal and performance they did not have an unambiguous means of ordering the conditions in the between subjects design they used. Gilliland (1980), in a between groups design with three levels of caffeine did find a curvilinear (inverted U) relationship between caffeine dose and GRE performance for low impulsives and a monotonically increasing function for the high impulsives. Stronger evidence has been reported by Anderson (1990) who, in a within subjects design with multiple levels of caffeine, found a reliable number of subjects showing an inverted U relationship between GRE performance and caffeine. More